This circuit, which enables us to track moving objects,
serves as an example of other brain circuits, some of which
perform thousands of computations every second. The findings
could aid the design of bionic eyes that track motion and
process visual information like our own eyes.
"This work reveals a very sophisticated neural computation,
the first non-linear computation performed by the nervous
system for which a circuit is close to being solved," said
Frank Werblin, professor of molecular and cell biology at
UC Berkeley. "It is a preliminary step in understanding
how more sophisticated computations are performed by the
brain."
Werblin notes, for example, that we use motion detection
every time we cross the street, anticipating when traffic
will reach our intersection and deciding when to cross.
"Barry Bonds probably has superior motion-detecting neurons,"
he added, referring to the home-run hitter with the San
Francisco Giants. "He takes a simple movement detector and,
in the context of a highly sophisticated action, uses it,
along with about a million other computations, both sensory
and motor, to make contact with the ball."
The technically demanding experiments measuring the output
of cells in the retina were conducted in Werblin's laboratory
by graduate students Shelley I. Fried of the School of Optometry's
Graduate Group in Vision Science and Thomas A. M�nch of
the Helen Wills Neuroscience Institute. All are members
of UC Berkeley's Health Sciences Initiative, a group of
hundreds of basic researchers banded together to tackle
some of the major health problems of the 21st century.
Horace Barlow was at UC Berkeley in 1965 when he and colleague
William Levick noticed that some cells in the retina of
rabbits fired only when a light moved through the eye's
field of view. A stationary light generated no response
from these cells except when blinking on or off.
In further experiments at UC Berkeley and, later, Cambridge
University, Barlow showed that some cells fire only when
a target moves from left to right, others fire only when
a target moves right to left, and still others respond only
to up-down or down-up movement. These cells activate the
four sets of muscles that control eye movement and allow
close tracking of moving objects. But the signals also make
their way through the optic nerve to the brain, providing
dynamic detail of the physical world.
Since these early experiments, scientists have discovered
cells in the retina that respond best to moving edges or
to moving edges of a particular orientation. There may be
a dozen or more types of these specialized ganglion cells.
The cells' axons - the outgoing wires of the neuron - bundle
together to form the optic nerve that funnels visual information
to the brain. Werblin and his laboratory colleagues have
been probing these cells in the retina, and building computer
models that help them understand how the physical world
is reconstructed by the eye and the brain into a picture
of our surroundings. Their findings have gone into the design
of a bionic eye that employs a unique computer chip that
can be programmed to do visual processing just like the
retina.
Barlow's original explanation of why directionally selective
cells fire only in response to movement in one direction
were very general, and recent experiments have confused
the picture even more. The UC Berkeley team has laid out
nearly the whole circuit, which includes three separate
and redundant mechanisms in the retina that work together
to create a directionally selective output.
"When Horace Barlow discovered that signals from some cells
specified the direction of movement, he tried to explain
it, and his explanation has been the reigning assumption,"
Werblin said "He was right in general outline, but his explanation
contained a lot of black boxes. We've shown what's going
on in the boxes."
The basic light detectors in the retina are the photoreceptors,
which fire off signals to a layer of horizontal cells and
thence to bipolar cells. The bipolar cells funnel signals
down their axons and relay them to the dendrites, or input
wires, of ganglion cells, which send the processed information
to the brain. All these cell types are arrayed in unique
layers, stacked one atop the other in the retina. At the
bottom of the stack are the 12 or so different kinds of
ganglion cells, including the directionally selective ganglion
cells.
If there were no other cells types, light shining on a
photoreceptor would initiate a signal that cascades unaltered
through the cell layers to the ganglion cell, and then into
the brain. But other cells, called amacrine cells, weave
among the bipolar cell axons and alter their output. This
is what makes some ganglion cells respond to motion in one
direction only.
Fried, M�nch and Werblin showed that the key player in
detecting motion is an amacrine cell dubbed a starburst
cell, because its dendrites spread out from the cell center
in a spoke-like pattern that suggests a figurative starburst.
The UC Berkeley team developed a technique to measure not
only the firing of the directionally selective ganglion
cells, but also the input these cells receive from starburst
and bipolar cells. In contrast to a recent paper suggesting
that starburst cells are not crucial to detecting directional
motion, they found them to be the critical link.
"The problem is that there are directionally selective
cells and starburst cells everywhere, and you can't tell
which are connected," M�nch said.
By probing this section of the retina, they discovered
that directionally sensitive cells are connected to only
half of the 60-70 starburst cells within their reach. All
of these are on one side only, an asymmetry that allows
them to veto firing when a stimulus comes from that direction.
Starburst cells not only affect directionally sensitive
ganglion cells, they also reach out and touch bipolar cells.
This allows starburst cells to alter the firing of bipolar
cells, too.
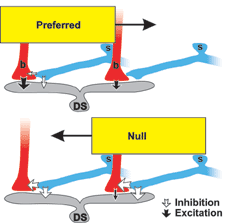
As a result, when a target moves in the "preferred" direction
- the direction that causes directionally sensitive ganglion
cells to fire - the ganglion cells get a strong stimulus
from bipolar cells and reduced inhibition from starburst
cells. The cumulative effect is a stimulus that triggers
the ganglion cell to send a signal to the brain.
When a target moves in the "null" direction, starburst
cells inhibit firing and also veto the excitation from bipolar
cells. The net effect is strong inhibition and no firing
of the directionally selective ganglion cells. Also, because
the inhibition from starburst cells arrives before any excitation
from the bipolar cells, firing is diminished even more.
"Starburst cells not only shut off the output of bipolar
cells, they also deliver negative input to the ganglion
cells," Fried said.
One unexpected finding was that starburst cells themselves
are sensitive to the direction of a moving stimulus. They
emit more neurotransmitter when a target moves outward along
a dendrite than when it moves inward toward the cell body,
creating a third mechanism by which they can signal direction
of movement.
"The original problem was, how do directionally selective
cells work," Werblin said. "Now we want to know how the
processes of starburst cells are selective."
The work was supported by grants from the Office of Naval
Research and the National Institutes of Health.